
Il parente più prossimo dell'antenato degli eucarioti sono gli Hodarchaeali, membri del superphylum Asgard, regno Archea
In un precedente articolo (in calce al presente) si è parlato dell’origine degli eucarioti e di come il gruppo di Archea che va sotto il nome di Asgard rappresenti (i discendenti oggi viventi del) l’anello di congiunzione tra eucarioti e protoeucarioti. Un’analisi complicata dal fatto che non esistono fossili di eucarioti più vecchi di 2 miliardi di anni fa, il che fa pensare che prima esistessero solo vari tipi di batteri.
È di qualche giorno fa un articolo che aggiunge un tassello alle nostre conoscenze con la ricostruzione filogenetica di questo “antenato”.
Nei dibattiti in corso sull'eucariogenesi - la serie di eventi evolutivi che portano all'emergere della cellula eucariotica da antenati procarioti - i membri dell'archaea di Asgard sono stati identificati come i parenti arcaici più stretti degli eucarioti. Rimane tuttavia poco compresa la natura e l'identità filogenetica dell'ultimo antenato comune tra “noi” e gli Asgard archae.
Un indizio viene da un articolo pubblicato su Nature in cui si riporta l’analisi di centinaia di genomi da vari ceppi di Asgard e il risultante profilo dell’antenato comune. Il risultato vede gli eucarioti collocati come un clade annidato all'interno di Asgard archaea e il “gemello” ancora esistente di quello da cui sono emanati gli eucarioti negli Hodarchaeali, un nuovo ordine proposto all'interno di Heimdallarchaeia.
 |
| L'albero filogenetico. Notare gli Eukarya e gli Hodarchaecales Image credit: Laura Eme et al, (2023) Nature |
Gli Hodarchaeali sono stati scoperti nei sedimenti dei fondali oceanici e presso diverse sorgenti termali. Ad oggi si è riuscito a coltivare in laboratorio due ceppi; dall’analisi genetica si è potuto inferire il loro metabolismo che suggerisce che l’antenato comune fosse un chemolitotrofio termofilo mentre quello da cui poi sarebbero sorti gli eucarioti si sia adattato a condizioni mesofile acquisendo anche il potenziale genetico per uno stile di vita eterotrofico.
Articolo successivo sul tema Il quasi citoscheletro degli Asgard
Fonte
- Inference and reconstruction of the heimdallarchaeial ancestry of eukaryotes
Laura Eme et al, (2023) Nature
Articoli precedenti sul tema
- Alla ricerca di LUCA (articolo copiato in calce)


Shirt e libro di testo per veri nerd (Amazon)
*** Alla ricerca di LUCA ***
(febbraio 2023)
Alla ricerca di LUCA (Last Universal Common Ancestor) si è trovato un organismo con caratteristiche simili al proto-eucariote
Ne è passata di acqua sotto i ponti dai tempi della classificazione linneana degli organismi (per non parlare di Aristotele) quando ancora la genetica era un non-concetto e mancavano quindi gli strumenti di tracciamento che andassero oltre alla mera comparazione funzionale e strutturale.
In tempi più recenti si era giunti ad una classificazione degli organismi in 2 domini (Monera e Eukaryota) che però si scoprì in fretta essere troppo semplicistica. Il dominio Monera era tutto fuorché uniforme e si giunse presto a capire che al suo interno erano raggruppati due domini tanto diversi tra loro quanto i Monera dagli eucarioti. Si deve a Carl Woese nel 1990 la ripartizione del primo nei due domini Bacteria e Archaea, che vanno così ad aggiungersi agli eucarioti.
A scompaginare quella che sembrava una classificazione “finale” fu il problema dell’origine degli eucarioti che, seguendo una ipotesi formulata negli anni 60 da Lynn Margulis, sarebbero il risultato di una relazione endosimbionte seriale tra un ipotetico proto-eucariote ed un batterio. Evento avvenuto in più riprese come l’esistenza stessa di plastidi (in piante e alghe) e mitocondri (in tutti gli eucarioti) dimostra e spiegabile unicamente con una simbiosi estrema tra il "predatore" e un batterio non digerito dotato di funzionalità fotosintetiche o di fosforilazione ossidativa basata sull’ossigeno, a seconda del tipo.
Se questo spiegava la presenza e le caratteristiche del DNA in questi organelli e la forte somiglianza metabolica e della sintesi proteica tra un Archea e un eucariote, rimaneva da spiegare il mistero del nucleo.
Questa la ragione per cui oggi si preferisce posizionare gli Archea come un "ramo" separatosi molto precocemente da un proto-eucariote invece che come un terzo "ramo".
Il nucleo, elemento caratterizzante la cellula eucariote è infatti dotato di doppia membrana come i plastidi e mitocondri; ma se la doppia membrana dei primi era spiegabile come “il residuo” del processo di fagocitosi avvenuto eoni fa, questa ipotesi perdeva di senso per il nucleo a meno di ipotizzare la cattura del genoma da un altro ipotetico antenato, divenuto poi dominante (idea francamente poco sostenibile).
 |
| La struttura di mitocondri e cloroplasti ha in comune l'essere dotata di doppia membrana (image credit: biologyexams4u) |
 |
| Il nucleo è dotato di una doppia membrana di cui quella esterna in continuità con il reticolo endoplasmico |
A dirimere il nodo alcuni articoli apparsi nell’ultimo biennio, capaci di spiegare sia l’evento di cattura dei batteri che sarebbero poi diventati i plastidi e i mitocondri, che la doppia membrana nucleare.
Punto centrale, l’identificazione di un Archea in cui si notano chiaramente lunghe protrusioni (simili agli pseudopi degli eucarioti) usate per catturare il cibo dall’ambiente, finora mai osservate negli archea.
Comprendo che il tema non sia semplice.
Questa la ragione per cui ho deciso di ripescare un vecchio articolo in cui trattavo di una classe di Archea con caratteristiche molto simili a quelle del proto-eucariote, modificandolo in modo sostanziale sia in queste righe introduttive che nella parte finale contenente i vari aggiornamenti. Il risultato, qui riproposto è un articolo nuovo, lungo ma unico con cui avere una panoramica (poco tecnica) dell'argomento. Nell'ultimo paragrafo ho inserito il riassunto degli argomenti toccati.
Nota. Il processo endosimbiontico è avvenuto più volte nel corso dell'evoluzione. Di seguito una figura riassuntiva
| Le piante sono il risultato di almeno 3 eventi di endosimbiosi |
***
Loki, Asgard e Midichlorian
Dai fumetti i nomi di batteri con ruolo chiave nell'evoluzione
Tutto nasce da un curioso intreccio tra biologia e Star Wars, citazione non casuale se si pensa ad un Archea che deve il suo nome ai Midi-chlorian, la fonte stessa della Forza nella narrazione dell'universo di Lucas) al cui articolo rimando prima di continuare questo.
Loki, Asgard e Midichlorian, i microbi allla base della Forza in Guerre Stellari
Usare questi nomi come incipit di un articolo potrebbe fare pensare al repentino reindirizzamento del blog verso tematiche comics in salsa Marvel o SF in genere. Non che sia immune a tale fascinazione ma … no, si rimane sempre in ambito biologia.
Usare questi nomi come incipit di un articolo potrebbe fare pensare al repentino reindirizzamento del blog verso tematiche comics in salsa Marvel o SF in genere. Non che sia immune a tale fascinazione ma … no, si rimane sempre in ambito biologia.
 |
| Non solo mitologia norrena ma anche nomi di batteri (credit: Uppsala University) |
I nomi evocativi di cui sopra sono stati infatti usati per identificare particolari microorganismi particolarmente interessanti da un punto di vista filogenetico (studio delle linee di discendenza nell'evoluzione della vita).
Se qualcuno avesse chiesto un secolo fa ad uno scienziato di fare una distinzione macro degli organismi viventi, questi avrebbe certamente usato quella tra animali e vegetali. Distinzione poi superata (piante e funghi hanno ben poco in comune) in favore di quella tra cellule prive di nucleo (procarioti o Monera cioè i batteri) e con nucleo (eucarioti).
Se qualcuno avesse chiesto un secolo fa ad uno scienziato di fare una distinzione macro degli organismi viventi, questi avrebbe certamente usato quella tra animali e vegetali. Distinzione poi superata (piante e funghi hanno ben poco in comune) in favore di quella tra cellule prive di nucleo (procarioti o Monera cioè i batteri) e con nucleo (eucarioti).
 |
| Una distinzione molto generica (e non più corretta) che sottintende organismi molto diversi tra loro |
Alla metà degli anni '70 del secolo scorso la tecnologia fornì strumenti di indagine innovativi che permise ai ricercatori, tra cui Carl Woese, di analizzare la sequenza del RNA ribosomale e di usarla per determinare i rapporti di parentela tra i vari organismi. L'analisi mostrò che i procarioti presentavano una eterogeneità tale che alcuni di loro erano più simili ad un eucariote (pur non avendo il nucleo) rispetto ai loro "consimili".
Si passò quindi, tra non poche discussioni, ad un albero della vita con 3 domini, due dei quali originati da quello che prima erano i procarioti: gli archeabatteri (oggi noti come Archea, batteri che vivono in ambienti estremi, che richiamano molto le condizioni della Terra primordiale); gli eubatteri (o semplicemente Bacteria, i batteri propriamente detti) e gli eucarioti (Eucarya, che comprendono i 5 regni Animalia, Fungi, Plantae, Protista, Chromista).
Lo scontro "ideologico" era tra il modello a tre domini di Woese e quello a due domini (vedi figura sotto).
Si passò quindi, tra non poche discussioni, ad un albero della vita con 3 domini, due dei quali originati da quello che prima erano i procarioti: gli archeabatteri (oggi noti come Archea, batteri che vivono in ambienti estremi, che richiamano molto le condizioni della Terra primordiale); gli eubatteri (o semplicemente Bacteria, i batteri propriamente detti) e gli eucarioti (Eucarya, che comprendono i 5 regni Animalia, Fungi, Plantae, Protista, Chromista).
Lo scontro "ideologico" era tra il modello a tre domini di Woese e quello a due domini (vedi figura sotto).
 |
| Il cladogramma in accordo con le osservazioni di Woese. Il cladogranma collega tutti gli organismi viventi al LUCA (il tronco nero nella parte inferiore). Questo grafico è derivato dalla sequenza del RNA ribosomico. (Image credit: Eric Gaba - NASA Astrobiology Institute) |
 |
| La differenza tra il modello proposto da Woese (in alto), detto a 3 domini, e quello proposto da altri (in basso) detto a 2 domini è nel dove collocare gli eucarioti (cioè noi): originati a un antenato comune con gli Archaea oppure "figli", cioè emersi da un sottogruppo di Archaea? (image credit: Crion via wikipedia) |
 |
| Una versione recente (2017) e dettagliata del modello a due domini della precedente figura (image credit: Laura Eme et al, Nat. Microb.) |
Domanda che sorge spontanea è cosa si intenda per LUCA.
La parola è l'acronimo di Last Universal Common Ancestor, un termine che non deve ovviamente essere preso al singolare (organismo unico) ma come comunità di quasi-organismi protocellulari da cui sarebbero sorte le cellule propriamente dette.
Cosa e come fosse questa protocellula o quello che c'era prima ancora è solo frutto di ipotesi. Probabile che la transizione chiave sia avvenuta con il passaggio da materiale genetico "libero" e dotato di attività catalitica (come possono essere i ribozimi) ad una protocellula in seguito all'inglobazione di tale materiale in una vescicola lipidica micellare, definendo così un ambiente "interno" da uno "esterno".
Scegliere il modello corretto tra quello a 2 o a 3 domini implica capire se gli eucarioti siano originati dagli archèobatteri (definibili come sottogruppo di essi) oppure da un proto-archèobatterio (quindi siano diversi).
Oltre a questo è necessario capire come e quando sia avvenuto il passaggio da una cellula senza nucleo (e senza organelli) ad una con nucleo e mitocondri (più cloroplasti, il tipo di plastidi più noti ai più, nel caso delle cellule vegetali).
Oltre a questo è necessario capire come e quando sia avvenuto il passaggio da una cellula senza nucleo (e senza organelli) ad una con nucleo e mitocondri (più cloroplasti, il tipo di plastidi più noti ai più, nel caso delle cellule vegetali).
Almeno per quello che riguarda mitocondri e cloroplasti la teoria oggi accettata è quella della endosimbiosi proposta a fine anni '60 da Lynn Margulis.
In estrema sintesi le attuali cellule eucariote sarebbero il prodotto di una simbiosi tra un protoeucariote anaerobio dotato di nucleo in cui è racchiuso il genoma e una cellula batterica aerobica, priva di nucleo, capace di utilizzare l'ossigeno; con il tempo quest'ultima si trasformò nella centralina energetica della cellula grazie alla sua capacità di coniugare la neutralizzazione dell'ossigeno con una elevata produzione di energia chimica (sotto forma di ATP) grazie alla fosforilazione ossidativa.
In verità più che simbiosi, l'evento sarebbe da attribuire ad un pasto "indigesto" con il quale il protoeucariote inglobò il batterio senza però riuscire a ucciderlo e a digerirlo, portandoselo appresso nelle generazioni successive. Negli eoni che seguirono il rapporto tra i due divenne così vantaggioso per entrambi da culminare in una migrazione di molti geni batterici nel genoma del nucleo (che quindi prese il controllo della "ex" cellula batterica). Questo "ex" batterio divenne il mitocondrio (ancora oggi ha un suo genoma "essenziale" ed è capace di divisione autonoma) ed è stato l'elemento discriminante che ha permesso alla cellula eucariote di "evolvere" fino agli organismi multicellulari: il possedere una propria centrale energetica (in numero fino al migliaio come negli spermatozoi) fornisce un chiaro vantaggio al possessore.
La simbiosi si è spinta talmente da essere oggi di fatto "obbligata". Tra gli eucarioti esiste solo un protista appartenente al genere Monocercomonoides che è privo di mitocondri. Negli organismi pluricellulari alcune cellule decidono di farne a meno semplicemente "perdendoli" durante il processo differenziativo; l'esempio classico sono gli eritrociti (globuli rossi) che si spingono talmente in là nel loro processo minimalista tutto dedito al trasporto di ossigeno che oltre ai mitocondri perdono perfino il nucleo, cioè il loro DNA. Per approfondimenti l'articolo --> "The origin of mitochondria" su Nature)
Ed ecco che arriviamo ai Midi-chlorian.
Il nome, coniato nell'episodio 1 di Guerre Stellari, indica gli esseri microscopici da cui "emana" la Forza (presente in tutti i viventi ma solo alcuni individui ne possiedono un numero così elevato da fare emergere questa "energia"). Chiaramente era solo un escamotage narrativo per spiegare un fenomeno che non voleva essere percepito come magico e come tale appartenente al regno del fantasy.
Fu solo in seguito, con la identificazione di alcuni batteri che avevano molte caratteristiche che richiamavano gli antenati dei mitocondri (cioè i batteri inglobati e sfuggiti al pasto) che si decise di usare questo nome nella scienza "ufficiale". Del resto quale nome migliore di quello da cui origina la "forza" per indicare il progenitore delle centraline energetiche delle cellule?
(per approfondimenti vedi --> "Dalla Forza l'ispirazione per il nome dell'antenato dei mitocondri")
Se il "fossile vivente", discendente della famiglia da cui originarono i mitocondri è oggi associato al Midichloria mitochondrii, mancava all'appello il discendente (sempre che esistano tuttora) del proto-eucariote che prima tentò di papparsi il batterio e poi finì per stringere con esso una alleanza, anzi una "dipendenza", mutualmente necessaria.
Il batterio più strettamente imparentato con l'antenato dei cloroplasti è invece il cianobatterio (capace di fotosintesi) Gloeomargarita lithophora. Il processo endosimbiontico sembra essere avvenuto altre volte, dopo quello fondante che ha originato le cellule vegetali circa 1,5 miliardi di anni fa; si parla di endosimbiosi secondaria o terziaria nota anche come kleptoplasty che letteralmente indica il rubare il plastidio da una fonte di cibo (eventi noti in dinoflagellati ad esempio).
L'organismo ricevente, cioè il proto-eucariote, era con ogni probabilità un Archea.
Ed eccoci arrivati ai nomi Loki e Asgard.
Tutto nasce con il sequenziamento del genoma dei microbi prelevati dai fanghi dei fondali marini della Groenlandia. L'analisi, fatta da Anja Spang e Thijs Ettema, rivelò la presenza di strani Archea, che per alcune loro caratteristiche fuorvianti furono chiamati Lokiarchaeota (Loki nella mitologia norrena è il Dio dell'inganno e della confusione) e classificati come proteoarchaeota e nel superphylum Asgard.
La loro caratterizzazione ha riacceso un dibattito che si pensava sopito tra i fautori della divisione a due domini e chi propendeva per quella a tre domini. I Lokiarchaeota, pur chiaramente archeabatteri possiedono infatti alcuni geni (175, poco più del 3%) coinvolti in importanti funzionalità caratteristiche degli eucarioti. Tra questi i geni per:
- i sistemi proteici di membrane intracellulari;
- i pori nucleari;
- la fagocitosi;
- la duplicazione del DNA;
- l'ubiquitina (il sistema di degradazione delle proteine)
- ... e altro ancora.
I dati sembrano spingere verso il modello a due domini e con esso il claim "siamo tutti Archaea" diventerebbe una caratteristica intrinseca a tutto ciò, noi compresi, che non è batterio e non è virus (dal protozoo all'elefante; nota i virus sono meglio definibili come quasi-organismi).
Dato che l'universo Marvel e quello di Lucas sono ben forniti, non stupitevi se in futuro sentirete in ambito scientifico altri nomi derivati dal mondo comics et similia.
 |
| Un altro modo per vedere graficamente l'evoluzione dal LUCA ai suoi discendenti (image credit: Maulucioni via wikipedia) Un cladogramma più accurato --> QUI |
***
Identificato il gruppo di Archea con caratteristiche "idonee" a rappresentare il proto-eucariote ancestrale il quadro era ben lungi dall'essere completo sia per l'assenza di caratteristiche essenziali per spiegare come fosse avvenuto il processo di cattura che avrebbe portato all'endosimbiosi che per l'impossibilità di spiegare la doppia membrana nucleare.
Nota. Quando scrivo dell'identificazione di organismi con caratteristiche compatibili a quelle dell'antenato ancestrale, questo non vuol che questi organismi coincidano con l'antenato. Il paragone più semplice si riferisce a noi e alle scimmie, nostri "cugini" prossimi che sono tuttavia ben diversi dall'antenato in comune che è scomparso nello stesso momento in cui hanno cominciato ad accumularsi le differenze tra le varie linee evolutive che poi avrebbero portato ai primati attuali.
Studiare gli Archea non è semplice di suo e la difficoltà si amplifica quando si vuole esplorare la consistenza della teoria dell'endosimbionte in questi organismi. La spiegazione è che questi organismi vivono in nicchie ecologiche talmente particolari (letteralmente da Terra primordiale) da rendere difficile sia riprodurre le condizioni esatte in laboratorio, trovando i nutrienti corretti, che trovare eventuali "partner" nella catena alimentare necessari alla loro sussistenza. Immaginate poi di dovere studiare al microscopio un organismo che si divide molto più lentamente di altri batteri (condizione derivante dal vivere in ambienti poveri di nutrienti e difficili) anche nelle condizioni per lui ottimali e di farlo in condizioni di sicurezza per l'operatore quando il "brodo di coltura" è ad alta temperatura, acido e con effluvi tossici.
Nonostante queste difficoltà, le pubblicazioni su questo gruppo di microbi sono quasi raddoppiate negli ultimi dieci anni.
I modelli per spiegare la transizione tra un gruppo di Archea primordiale e il proto-eucariote differiscono tra loro sul "timing" dell'evento, inteso in quanto questo organismo avesse già acquisito peculiarità strutturali eucariote tipo la presenza di membrane flessibili e compartimenti interni, PRIMA di catturare il batterio che sarebbe diventato il mitocondrio. Se alcuni ipotizzano che queste cellule avessero già "moderni" meccanismi di cattura del materiale esterno (fagocitosi), altri come Sven Gould credono che questo sistema estremamente specializzato si sia sviluppato eoni dopo l'endosimbiosi, e che la cattura sia avvenuta mediante meccanismi più semplici della fagocitosi tipo quelli usate dai batteri.
Ma c'è un terzo modello che chiamiamo modello di Baum dal nome dei proponenti, due cugini.
Il modello di Baum si distingue dagli altri perché non solo spiega la cattura del "proto-mitocondrio" senza la fagocitosi ma anche l'origine della membrana nucleare.
L'idea nasce nel lontano 1984 quando uno dei due era ancora uno studente, formalizzata nel 2014. La riassumo di seguito.
Il processo inizia in un mondo in cui Archea e batteri condividono l'ambiente e con esso le risorse. Si ipotizza la presenza di un gruppo di Archea dotato di una sufficiente plasticità della membrana, capace di estenderne propaggini verso l'esterno in modo da aumentare la superficie, così da massimizzare lo scambio di nutrienti. Durante queste attività alcune propaggini (lunghe e più voluminose del "corpo cellulare") potrebbero avere circondato in modo casuale un più piccolo batterio fino a intrappolarlo in qualcosa di simile ad un endosoma. Un processo simile potrebbe essere avvenuto con il ritorno della propaggine verso il corpo cellulare (in cui di fatto si trovava confinato il genoma) avvolgendolo; a questo fenomeno sarebbero seguiti la fusione e riarrangiamento delle membrane con il risultato finale di una cellula di maggiori dimensioni, dotata di una membrana nucleare e di un sistema di membrane a delimitare compartimenti interni (immagine a destra della figura sottostante).
Un "percorso" ben diverso da quello associato al modello della fagocitosi, in quanto inizia ipotizzando l'esistenza di una comunità di organismi e in tempi molto più lunghi invece che con un singolo passaggio.
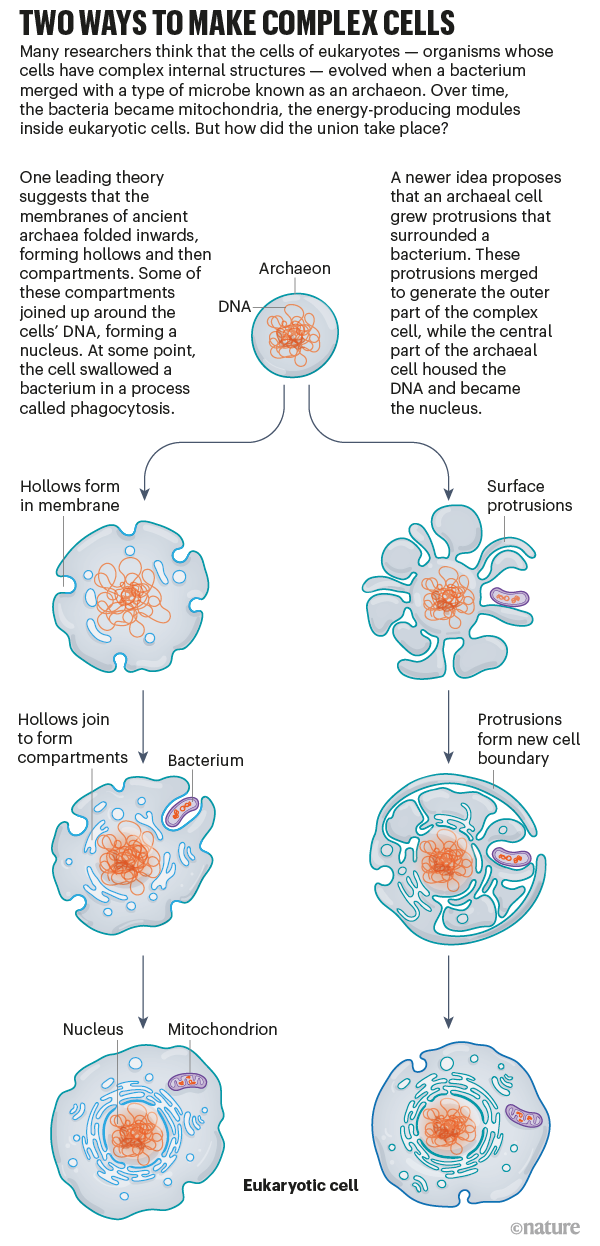 |
| A sinistra il modello classico basato sulla fagocitosi, a destra il modello dei Baum (image credit: B. Baum & D.A. Baum BMC Biol. via Nature) |
Modello affascinante e che riscosse una entusiastica accoglienza tra gli addetti ai lavori ma che mancava di prove sul campo, del tipo trovare un Archea con caratteristiche metaboliche compatibili e dotati di protrusioni di membrana sufficientemente ampie da adempiere allo scopo.
Un tassello che è arrivato nell'ultimo biennio con tre articoli, due centrati sullo studio di proteine essenziali per la divisione cellulare (Pulschen, A. A. et al. Curr. Biol. 2020 e Liao, Y. et al. Nature Microbiol, 2021) e l'altro con la caratterizzazione di un Archea cresciuto per la prima volta in laboratorio dotato di queste protrusioni (Imachi, H. et al. Nature, 2020).
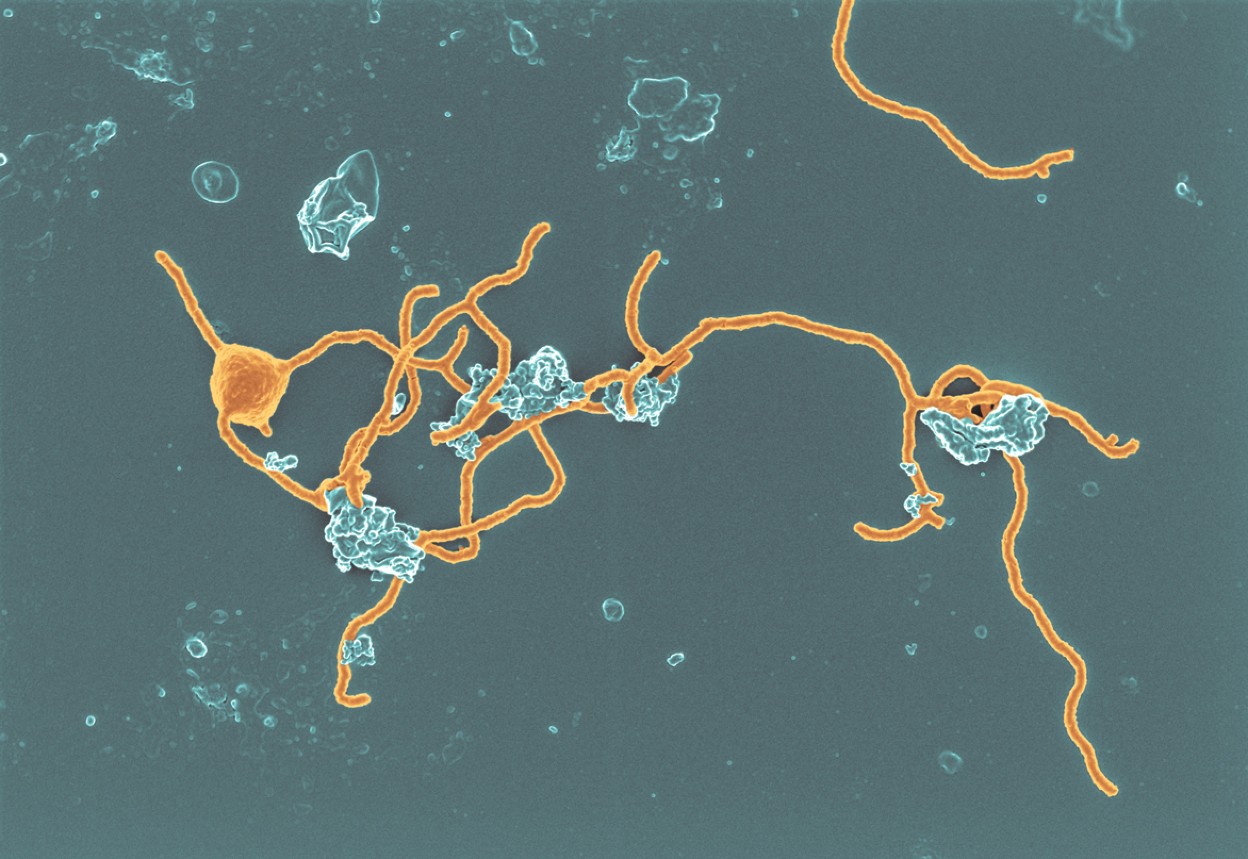
Cominciamo dal lavoro del 2020 in cui viene mostrato un Archea con protrusioni, dal nome temporaneo Prometheoarchaeum syntrophicum, appartenente al gruppo prima citato degli Asgard.
|
| Ci sono voluti 12 anni per riuscire a far crescere questo l'archea della foto, ritenuto simile al protoeucariote ancestrale. Image credit: Hiroyuki Imachi, Masaru K. Nobu, JAMSTEC via Nature |
Un Archea il cui tentativo di coltivazione è andato avanti per 12 anni fino a trovare condizioni giuste anche grazie all'aggiunta di alcuni batteri parte della stessa nicchia. La grande sorpresa arrivò solo quando le cellule smisero di dividersi con la comparsa di protrusioni molto simili a tentacoli usate per scandagliare l'ambiente e recuperare sostanze nutritive, ivi compresa l'interazione con altri batteri.
Una fotografia che entusiasmò i Baum perché spostava il loro modello dal limbo delle ipotesi affascinanti ma non verificabili, al reale.
I due articoli genetico-funzionali, uno dei quali portato avanti dai Baum, forniscono una evidenza genetica basata sull'evoluzione funzionale di proteine tra Archea ed eucarioti.
Lo studio nasce dalla premessa che il proto-eucariote altro non fosse che un Archea "modificato"; concetto oramai consolidato e ben evidente dal posizionamento del "ramo" eucariote sul ramo degli Archea, quindi dopo la separazione tra Bacteria e Archea.
Per trovare prove dell'evento l'unica possibilità era risalire l'albero evolutivo usando una qualche proteina funzionalmente essenziale (e il suo gene) come tracciante.
Una proteina adatta all'uopo deve essere parte di un sistema condiviso da tutte le cellule, qualunque sia il loro dominio di appartenenza. L'unico tratto funzionale condiviso da tutti è la creazione di copie di se stessi, cioè la divisione cellulare.
Lo studio si è focalizzato su due proteine presenti in tutti i microbi (FtsZ e SepF), essenziali per la creazione dell'anello divisorio tra le due cellule figlie, e negli eucarioti di proteine appartenenti alla classe ESCRT, coinvolte nella formazione di vescicole a partire dalla membrana plasmatica.
Gli Archea usati nello studio sono Haloferax volcanii (amante delle condizioni salmastre, come quelle del Mar Morto; il nome volcanii è fuorviante e deriva dallo scopritore, il microbiologo Benjamin Elazari Volcani) per FtsZ e SepF , e Sulfolobus acidocaldarius (qui il nome calza bene perché ama l'acido e il calore) per la proteina simile a ESCRT.
Lo studio in S. acidocaldarius evidenziò proprio proteine simili alle ESCRT usate però dall'Archea come componente dell'anello divisorio invece che nella formazione di vescicole. I dati suggerivano che quella fosse la funzione primigenia e che il differenziamento funzionale sia avvenuto negli eucarioti per gestire la maggiore complessità dei compartimenti interni assenti sia negli Archea che nei batteri.
I dati ottenuti hanno permesso di ipotizzare che in alcuni Archea ad un certo punto dell'evoluzione, le proteine fondamentali per la divisione cellulare sono andate incontro ad una rimodulazione funzionale passando dall'anello divisorio alle funzioni tipiche delle ESCRT oppure a compiti strutturali divenendo parte del citoscheletro (dalla FtsZ hanno avuto origine le tubuline, fondamentali per cellule strutturalmente e morfologicamente più complesse)
L'insieme di questi dati ha dato linfa vitale alla bella teoria dei Baum tanto che perfino manuali di riferimento classici come Molecular Biology Of The Cell pensano di sostituire la teoria della fagocitosi con questo modello.
In realtà c'è ancora tanta strada da fare per spiegare la formazione del complesso sistema di compartimenti membranosi interni. Una teoria in proposito viene da Sven Gould (fautore della teoria della cattura dei proto-mitocondri prima che gli Archea sviluppassero capacità di estroflettere) secondo la quale, data la capacità dei batteri e dei mitocondri di emettere vescicole è stata questa loro capacità ad avere reso la membrana degli eucarioti più simile a quella dei Bacteria invece che a quella, attesa se si da credito alla fagocitosi, degli Archea.
Riassumiamo i concetti chiave
- Gli eucarioti sono con ogni probabilità degli "Archea con upgrade" o in altre parole siamo più vicini evolutivamente agli Archea che non ai batteri (e lo stesso vale per il rapporto Archea-Bacteria)
- Gli Archea oggi esistenti più simili al proto-eucariote di partenza appartengono ad un gruppo noto come Asgard.
- Tra gli upgrade fondamentali quello chiave è consistito in uno o più endosimbiosi. Tra questi quello che ha permesso la comparsa del mitocondrio è comune a tutte le cellule eucariote, mentre quello/i associati ai plastidi sono unici per le cellule vegetali e alghe unicellulari.
- Il parente più prossimo dell'antenato del mitocondrio è il batterio Midichloria mitochondrii.
- Il parente più prossimo dei cloroplasti è il cianobatterio Gloeomargarita lithophora.
- La teoria endosimbiontica finora più accreditata era quella che prevedeva un evento di fagocitosi "fallito" come evento iniziale. La nuova teoria proposta dai Baum sta rapidamente guadagnando posizioni in quanto riesce a spiegare vari eventi altrimenti di origine ignota come la presenza di una doppia membrana nucleare.
- A supporto di questa teoria 3 studi che hanno mostrato l'esistenza di lunghe estroflessioni in almeno un Archea, e l'evoluzione dei geni necessari per la separazione delle cellule durante la divisione cellulare in geni utilizzati per funzioni complesse tipiche delle cellule eucariote.
Articolo associato al tema qui discusso --> "Una endosimbiosi come quella ancestrale, sta avvenendo anche ora"
- Asgard archaea illuminate the origin of eukaryotic cellular complexity
A. Spang, TJ Etterna et al, Nature. 2017;541(7637):353-358
- Proposal of the reverse flow model for the origin of the eukaryotic cell based on comparative analyses of Asgard archaeal metabolism.
A. Spang et al, Nat Microbiol. 2019;4(7):1138-1148
- Genomes of Asgard archaea encode profilins that regulate actin
Caner Akıl & Robert C. Robinson. Nature (2018), 562, pp. 439–443
- Proposal of the reverse flow model for the origin of the eukaryotic cell based on comparative analyses of Asgard archaeal metabolism.
A. Spang et al, Nat Microbiol. 2019;4(7):1138-1148
- Genomes of Asgard archaea encode profilins that regulate actin
Caner Akıl & Robert C. Robinson. Nature (2018), 562, pp. 439–443
Uppsala universitet / news
- Imachi, H. et al. Nature 577, 519–525 (2020)
- Baum, D. A. & Baum, B. BMC Biol. 12, 76 (2014)
- Liao, Y. et al. Nature Microbiol. 6, 594–605 (2021)
- Nußbaum, P. et al, Preprint at bioRxiv (2020)
- Pende, N. et al. Preprint at bioRxiv (2020)
- Pulschen, A. A. et al. Curr. Biol. 30, 2852–2859 (2020)
- Sagan, L. J. Theor. Biol. 14, 255–274 (1967)
- Spang, A. et al. Nature 521, 173–179 (2015)
- Gould, S. B. et al, Trends Microbiol. 24, 525–534 (2016)

Nessun commento:
Posta un commento