[Aggiornato Febbraio 2023. Vedi in fondo all'articolo]
Per quanto possa sembrare strano, l'esistenza o meno del pianeta 9 continua a rimanere un mistero (vedi in proposito gli articoli precedenti sul tema). Gli astronomi sono riusciti a visualizzare il buco nero supermassiccio al centro della galassia M87 ma ci sono ancora molte domande inevase sul nostro "vicinato".
 |
| Il pianeta 9 (sempre che esista) Image credit: Caltech/R. Hurt (IPAC) |
Le ragioni tecniche sono comprensibili, in primis la differenza dimensionale e di "attività" dell'oggetto da visualizzare.
Per quanto distante 55 milioni di anni luce, M87* risulta più facile da studiare di Sgr A*, il buco nero della nostra galassia distante "solo" 26 mila anni luce, in quanto 1000 volte più massiccio e (dal nostro punto di osservazione) meno occultato dalle altre stelle della galassia. Inoltre mentre Sgr A* è tutto sommato "tranquillo", la "voracità" di M87* (e con esso i segnali che emette) lo rende più facile da studiare.
Se trasponiamo il tutto alla ricerca di un pianeta ipotetico, privo di attività energetiche proprie e troppo lontano per potere riflettere la luce solare, si potrà meglio comprendere la difficoltà di trovare (qualora esistesse per certo) il pianeta anche usando telescopi potenti come PanSTARRS e LSST.
La maggior parte degli oggetti del sistema solare sono visti grazie alla luce (nella gamma del visibile) riflessa che arriva loro dal Sole. Come ben noto la luce che un oggetto riceve è inversamente proporzionale alla distanza al quadrato dalla sorgente radiante; di questa solo una parte viene riflessa (parametro legato all'albedo) e ancora meno quella che raggiunge i telescopi terrestri.Nelle zone più esterne del sistema solare, la luce riflessa è probabile che rientri nella gamma dell'infrarosso, per cui servono telescopi adatti. Ad oggi tuttavia anche strumenti dedicati come il Wide-field Infrared Explorer (WISE) non ha dato i risultati sperati.
Plutone visto con il LORRI NASA / JHU-APL / Southwest Research Inst

Il nome del pianeta da trovare è quanto di più letterale si possa immaginare.
Dopo la retrocessione di Plutone a pianeta nano nel 2006 (primo esempio di una categoria che oggi annovera Eris, Igea, Sedna, Quaoar e Cerere) i pianeti propriamente detti rimasti erano 8, per cui il pianeta fantasma doveva essere il "pianeta 9". Pianeta messo nel cesto delle possibilità per spiegare alcune anomalie nell'orbita di Nettuno; oggi si sa che tali anomalie erano frutto di una errata valutazione della sua massa ma il tema permane per un anomalo allineamento di alcuni TNO (corpi transnettuniani) per spiegare manca una massa pari a 5-10 volte la Terra che deve trovarsi da qualche parte prima della gigantesca regione coperta dalla nube di Oort, in una orbita distante 400-800 UA dal Sole.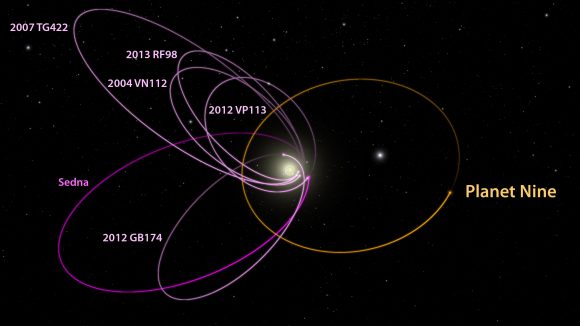
Altra possibilità è che la massa mancante sia sotto forma di piccoli oggetti sullo stile della fascia di asteroidi presente tra Marte e Giove.
Un nuovo tentativo è stato fatto usando l'Atacama Cosmology Telescope (ACT) capace di rilevare lunghezze d'onda millimetriche.
Il telescopio ha invero come scopo principale lo studio della radiazione cosmica di fondo (microonde), ma è ugualmente utile grazie alla sua alta risoluzione angolare.
Gli astronomi hanno ad oggi scansionato circa l'87% della volta celeste accessibile dall'emisfero australe per un periodo di sei anni, e i dati raccolti sono stati elaborati usando varie tecniche per massimizzare la probabilità di rilevare segnali deboli, anche se a scapito di informazioni sulla loro posizione esatta. Dall'analisi sono emersi circa 3500 candidati, nessuno delle quali però confermato dalle controanalisi.
In un risultato di fatto negativo si può anche vedere il bicchiere mezzo pieno: gli astronomi possono ora escludere con una sicurezza del 95% che vi sia un pianeta con le caratteristiche dimensionali cercate all'interno dell'area esaminata.
Per avere un quadro completo bisognerà attendere l'analisi della volta boreale (possibile solo con altro telescopio) e le analisi con telescopi specializzati per lunghezze d'onda superiori.
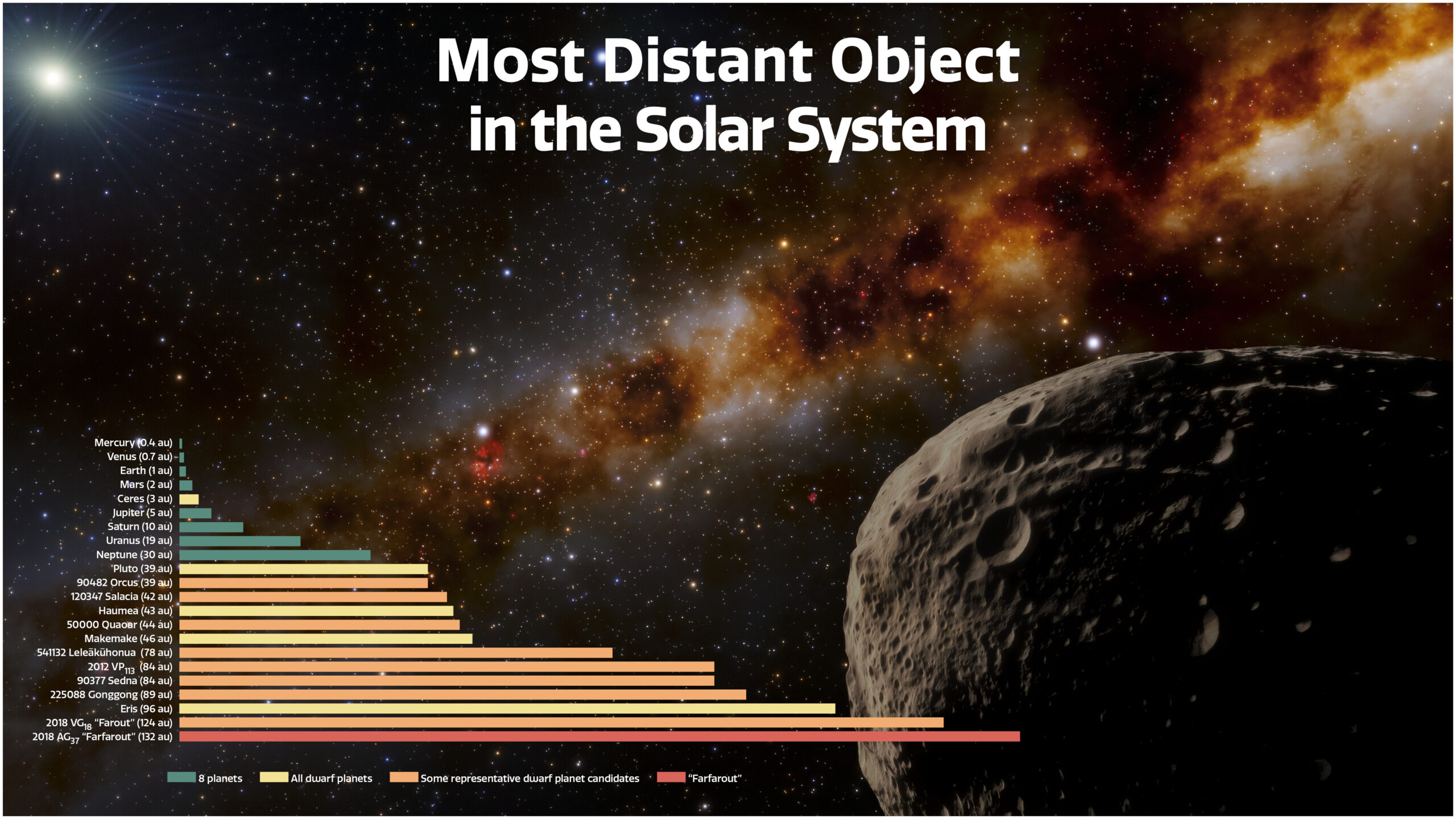 |
| Una panoramica dei corpi planetari principali del sistema solare. Ad oggi il più distante osservato è FarFarOut (traducibile come crasi dal significato "esternolontanolontano") Image credit: INAF |
Di seguito un video prodotto dalla Caltech di qualche anno fa sulla ricerca del Pianeta 9
AGGIORNAMENTO
Un nuovo approccio alla ricerca del Pianeta 9 arriva da un articolo da poco pubblicato su Astrophysical Journal da Man Ho Chan dell'Università di Hong Kong. La novità di questo metodo sta nel focalizzarsi sulle (potenziali) lune che molto verosimilmente il pianeta dovrebbe avere (presenti perfino intorno a planetoidi).
Plutone, l’ex pianeta retrocesso a pianeta nano e membro effettivo dei corpi della fascia di Kuiper, ha ben 5 lune,
Apparentemente un controsenso il cercare satelliti (per definizione di massa inferiore) di un pianeta “fantasma”, ma Chan spiega che se questi fossero presenti si generebbe una traccia di calore fluttuante dovuta al riscaldamento mareale, di entità 2,5 volte superiore a quelle dal pianeta e (potenzialmente) rilevabili dall’tacama Large Millimeter/submillimeter Array Observatory (ALMA).
Ad oggi la probabilità che le orbite anomale di molti corpi della fascia di Kuiper non sia dovuta al pianeta misterioso sono comprese tra lo 0,02% e lo 0,04%. Sempre che ovviamente la "massa mancante" non sia un buco nero primordiale le cui dimensioni minime (starebbe dentro uno zaino) renderebbe la sua identificazione quasi impossibile con gli strumenti oggi disponibili … a meno che non abbia anche lui dei satelliti da cui rilevare il calore mareale irradiato.
La ricerca al confini del sistema solare continua
Fonte
- The Atacama Cosmology Telescope: A Search for Planet 9
Sigurd Naess et al, The Astrophysical Journal, (dec 2021) 923 (2)
- Evidence for a Distant Giant Planet in the Solar System
K. Batygin and M.E. Brown, (2016) The Astronomical Journal, 151:22 (12pp)
- What if planet 9 has satellites?
Man Ho Chan, arXiv, Gennaio 2023
https://phys.org/news/2022-03-planet.html